65 The Cerebellum
Parts of the Cerebellum
The cerebellum, which looks like a separate structure attached to the bottom of the brain, plays an important role in motor control.
Learning Objectives
Describe the parts of the cerebellum
Key Takeaways
Key Points
- The cerebellum can be separated into three lobes: the flocculonodular lobe, anterior lobe, and posterior lobe.
- The medial zone of the anterior and posterior lobes constitutes the spinocerebellum, or paleocerebellum.
- There are about 3.6 times as many neurons in
the cerebellum as in the neocortex. - Based on surface appearance, three lobes can be distinguished in the
cerebellum: the flocculonodular lobe, anterior lobe (above the primary
fissure), and the posterior lobe (below the primary fissure).
Key Terms
- granule cells: An extremely small type of neuron that is the the smallest cell found in the brain.
- neocortex: The largest part of the cerebral cortex of the human brain, covering the two cerebral hemispheres.
- Purkinje cells: A class of GABAergic neurons located in the cerebellar cortex that are some of the largest neurons in the human brain.
GABA is the chief inhibitory neurotransmitter
in the mammalian
central nervous system. - cerebellum: Part of the hindbrain. In humans, it lies between the brainstem and the cerebrum. It plays an important role in sensory perception, motor output, balance, and posture.
The cerebellum has the appearance of a separate structure attached to the bottom of the brain, tucked underneath the cerebral hemispheres. The surface of the cerebellum is covered with finely spaced parallel grooves, in striking contrast to the broad irregular convolutions of the cerebral cortex.

Cerebellum: Brain section showing cerebellum position and structure
These parallel grooves conceal the fact that the cerebellum is actually a continuous thin layer of tissue (the cerebellar cortex), tightly folded in the style of an accordion. This thin layer contains several types of neurons with a highly regular arrangement, most importantly Purkinje cells and granule cells. This complex neural network gives rise to a massive signal-processing capability, but almost all of its output is directed to a set of small deep cerebellar nuclei lying in the interior of the cerebellum.
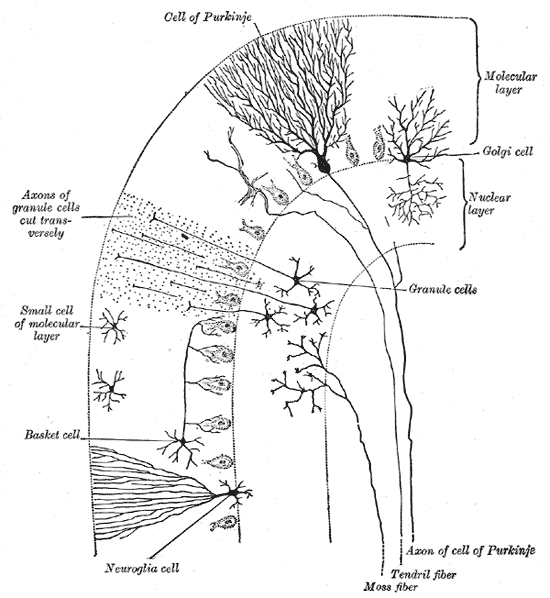
Cells of the Cerebellum: Transverse section of a cerebellar folium, showing principal cell types and connections.
The cerebellum is separated from the overlying cerebrum by a layer of leathery dura mater. Anatomists classify the cerebellum as part of the metencephalon, which also includes the pons, and all its connections with other parts of the brain travel through the pons. The metencephalon is the upper part of the rhombencephalon, or hindbrain. Like the cerebral cortex, the cerebellum is divided into two hemispheres. It also contains a narrow midline zone called the vermis. A set of large folds is, by convention, used to divide the overall structure into 10 smaller lobules. Due to its large number of tiny granule cells, the cerebellum contains more neurons than the rest of the brain put together, but comprises only 10% of total brain volume.
Based on surface appearance, three lobes can be distinguished in the cerebellum: the flocculonodular lobe, anterior lobe (above the primary fissure), and the posterior lobe (below the primary fissure). Excluding the flocculonodular lobe, which has distinct connections and functions, the cerebellum can be parsed functionally into a medial sector called the spinocerebellum and a larger lateral sector called the cerebrocerebellum. A narrow strip of protruding tissue along the midline is called the vermis (Latin for “worm”).

Divisions of the cerebellum: Schematic representation of the major anatomical subdivisions of the cerebellum. Superior view of an “unrolled” cerebellum, placing the vermis in one plane.
The smallest region, the flocculonodular lobe, is often called the vestibulocerebellum. It is the oldest part of the brain in evolutionary terms (archicerebellum) and participates mainly in balance and spatial orientation. Its primary connections are with the vestibular nuclei, although it also receives visual and other sensory input.
The medial zone of the anterior and posterior lobes constitutes the spinocerebellum, also known as the paleocerebellum. It receives proprioception input from the dorsal columns of the spinal cord (including the spinocerebellar tract) and from the trigeminal nerve, as well as from visual and auditory systems. It sends fibers to deep cerebellar nuclei that in turn project to both the cerebral cortex and the brain stem, thus providing modulation of descending motor systems.
The lateral zone, which in humans is by far the largest part, constitutes the cerebrocerebellum, also known as the neocerebellum. It receives input exclusively from the cerebral cortex (especially the parietal lobe) via the pontine nuclei (forming corticopontocerebellar pathways), and sends output mainly to the ventrolateral thalamus (in turn connected to motor areas of the premotor cortex and primary motor area of the cerebral cortex) and to the red nucleus.
Functions of the Cerebellum
Cerebellar function was once believed to be motor-specific, but newer findings suggest the cerebellum is also involved in higher-level brain processing.
Learning Objectives
Describe the function of the cerebellum
Key Takeaways
Key Points
- The cerebellum is essential for making fine adjustments to motor actions.
Cerebellar dysfunction primarily results in problems with motor control. - Four principles are important to cerebellar processing: feedforward processing, divergence and convergence, modularity, and plasticity.
- Signal processing in the cerebellum is almost entirely feedforward. Signals move through the system from input to output with very little internal transmission.
- The cerebellum both receives input and transmits output via a limited number of cells.
- The cerebellar system is divided into thousands of independent modules with similar structure.
Key Terms
- neural divergence: When a neuron fires and the signal is sent to many other neurons.
- neuroplasticity: Changes in neural pathways and synapses due to changes in behavior, environment, neural processes, or bodily injury.
- feedforward processing: A property of some neural circuits where signals move unidirectionally through the system from input to output with very little recurrent internal transmission.
Examining the consequences of damage to the the cerebellum provides the strongest clues to its function. Animals and humans with cerebellar dysfunction show problems with motor control. They can still generate motor activity, but lose precision and produce erratic, uncoordinated, or incorrectly timed movements. Functional imaging studies have also shown cerebellar activation in relation to language, attention, and mental imagery. Additionally, correlation studies have shown interactions between the cerebellum and non-motor areas of the cerebral cortex.
Principles of Cerebellar Function
The comparative simplicity and regularity of the cerebellar anatomy led to an early hope that a similar simplicity of computational function could be implied. This was expressed in one of the first books on cerebellar electrophysiology, The Cerebellum as a Neuronal Machine by John C. Eccles, Masao Ito, and Janos Szentágothai. Although a full understanding of cerebellar function has remained elusive, at least four principles have been identified as important: feedforward processing, divergence and convergence, modularity, and plasticity.
- Feedforward processing: The cerebellum differs from other parts of the brain (especially the cerebral cortex) in that the signal processing is almost entirely feedforward. This means signals move unidirectionally through the system from input to output with very little recurrent internal transmission. The small amount of recurrence that does exist consists of mutual inhibition. There are no mutually excitatory circuits. This feedforward mode of operation means that the cerebellum, in contrast to the cerebral cortex, cannot generate self-sustaining patterns of neural activity. Signals enter the circuit, are processed by each stage in sequential order, and then leave. This provides a quick, concise response to any combination of inputs.
- Divergence and convergence: In the human cerebellum, information from 200 million mossy fiber inputs is expanded to 40 billion granule cells. This neural divergence is followed by parallel fiber outputs that converge onto 15 million Purkinje cells. Due to their longitudinal alignment, the approximately 1000 Purkinje cells belonging to a microzone may receive input via neural convergence from as many as 100 million parallel fibers. The cells then focus their own output down to a group of less than 50 deep nuclear cells. Therefore, the cerebellar network only receives a modest number of inputs to process and send results via a limited number of output cells.
- Modularity: The cerebellar system is functionally divided into thousands of independent modules. All modules have a similar internal structure but different inputs and outputs. A module consists of a small cluster of neurons in the inferior olivary nucleus, a set of long narrow strips of Purkinje cells in the cerebellar cortex (microzones), and a small cluster of neurons in one of the deep cerebellar nuclei. Different modules share input, but also appear to function independently. The output of one module does not seem to significantly influence the activity of other modules.
- Plasticity: The synapses between parallel fibers and Purkinje cells and between mossy fibers and deep nuclear cells are both susceptible to modification of their strength. In a single cerebellar module, input from as many as a billion parallel fibers converge onto a group of less than 50 deep nuclear cells, and the influence of each parallel fiber on those nuclear cells is adjustable. This arrangement gives tremendous flexibility for fine-tuning the relationship between cerebellar inputs and outputs.
Role of the Cerebellum in Motor Learning
There is considerable evidence that the cerebellum plays an essential role in some types of motor learning, most clearly in tasks in which fine adjustments must be made to an action’s performance. There has been much dispute about whether learning takes place within the cerebellum itself, or whether it merely serves to provide signals that promote learning in other brain structures.
One of the most extensively studied cerebellar learning tasks is the eyeblink conditioning paradigm. A blink response is elicited when a neutral conditioned stimulus, such as a tone or a light, is repeatedly paired with an unconditioned stimulus, such as an air puff.
After many conditioned-unconditioned stimuli (CS-US) pairings, an association is formed whereby a learned blink, or conditioned response, occurs and precedes US onset. The magnitude of learning is measured by the percentage of all paired CS-US trials that result in a CR.
Experiments showed that lesions localized either to a specific part of the interpositus nucleus (one of the deep cerebellar nuclei), or to a few specific points in the cerebellar cortex, abolished learning of a correctly timed blink response. If cerebellar outputs are pharmacologically inactivated while leaving the inputs and intracellular circuits intact, learning takes place even while the animal fails to show any response. However, if intracerebellar circuits are disrupted, no learning takes place; these facts taken together make a strong case that learning occurs inside the cerebellum and that its cells exhibit neuroplasticity.
Clinical Relevance
Motor abnormalities are the primary symptoms of cerebellar dysfunction, and the nature of those depends on the part of the cerebellum affected.
Example Symptoms of Cerebellar Dysfunction
- Damage to the flocculonodular lobe (vestibulocerebellum): loss of equilibrium causing an altered
walking gait - Lateral zone damage: problems with skilled voluntary and planned movements leading to errors in intended movements (eg., dysdiadochokinesia, the inability to perform rapid
alternating movements). - Damage to the midline portion: disruption of whole-body movements
- Damage to the upper part of the cerebellum: gait impairments and other problems with
leg coordination (ie, ataxia).
